
Samenvatting
- Apoptose, fysiologische celdood, speelt een rol in de pathogenese van veel ziekten, met name van maligniteiten, degeneratieve aandoeningen en ouderdomsziekten.
- De kennis over het mechanisme van de apoptose biedt de farmaceutische industrie de uitdaging nieuwe stoffen te ontwikkelen, gericht tegen de specifieke afwijkingen in de zieke weefsels en het is aan de klinische geneeskunde om deze middelen op hun therapeutisch potentieel te testen.
- Geneesmiddelen die momenteel worden onderzocht, zijn gericht op modulatie van de regulatoren van apoptose, de intracellulaire signaaloverdracht, de productie van groeifactoren en de expressie van receptoren. Daarbij gaat het bijvoorbeeld om gentherapie en de toepassing van antisense DNA-fragmenten.
- De recente resultaten verkregen bij behandeling van kanker zijn veelbelovend en laten ruimte om deze nieuwe benadering te combineren met traditionele cytostatische geneesmiddelen.
- Het is te verwachten dat in de komende jaren ook een biologisch-therapeutische benadering van degeneratieve ziekten, waarbij de apoptose moet worden afgeremd, tot ontwikkeling zal komen.
artikel
Het bestaan van een fysiologische vorm van celdood werd voor het eerst geopperd in 1972 door Kerr et al.1 Zij noemden deze vorm van celdood ‘apoptose’ ter onderscheid van necrose, celversterf dat optreedt na een letale beschadiging. Zij veronderstelden dat in het genoom een programma zit dat de apoptose bestuurt. Ook reeds tijdens de embryonale ontwikkeling treedt celeliminatie op, aangeduid als ‘geprogrammeerde celdood’. In de literatuur worden beide termen door elkaar gebruikt. Om verwarring te vermijden gebruiken wij hier apoptose.
Aanvankelijk bestond bij veel biologen grote scepsis over het bestaan van een genetisch celdoodprogramma.2 Van die aanvankelijke twijfel is door de huidige kennis over het moleculair-biologisch mechanisme thans geen sprake meer. Apoptose blijkt tijdens de evolutie vanaf de primitiefste organismen tot bij de mens toe in het genoom geconserveerd te zijn gebleven.3 Al snel zag men in dat manipulatie van apoptose een machtig middel kan zijn bij de bestrijding van aandoeningen waarbij dit systeem ontspoord is. In de wetenschappelijke en de farmaceutische wereld werd daarom veel onderzoek en geld geïnvesteerd om het mechanisme van de apoptose in kaart te brengen.
Tot op heden heeft apoptose bij de praktiserende arts weinig belangstelling gewekt. Dit is begrijpelijk wanneer men bedenkt dat de nieuwe kennis over de apoptose tot nu geen bruikbare aanvulling heeft geleverd aan het medisch-diagnostisch of therapeutisch arsenaal. De laatste jaren is dat aan het veranderen. Het is duidelijk geworden dat bepaling van de intracellulaire eiwitten die betrokken zijn bij het apoptosemechanisme, informatie kan verschaffen over de oorzaak van een aantal ziekten en over de prognose ervan. Nog sterker, met genmanipulatie is het mogelijk gebleken bij sommige ziekten de neiging tot apoptose zo te moduleren dat er therapeutische resultaten werden geboekt.
In dit artikel beschrijven wij de kennis van de celbiologie van apoptose en de eerste behandelingsresultaten verkregen met het moduleren van het apoptotisch mechanisme. Het is te verwachten dat de komende jaren apoptose een toenemende betekenis zal krijgen voor de diagnostiek en de therapie van een aantal tot nu onbehandelbare ziekten.
het moleculair mechanisme
Hier beperken wij ons tot die aspecten die nodig zijn om het mechanisme van apoptose te kunnen toelichten. Gedetailleerde informatie over de biochemie van apoptose kan men vinden in recente overzichtsartikelen.4-9 De figuur laat zien dat het proces van de apoptose in 3 fasen kan worden onderscheiden.
De inductiefase
De inductie van apoptose kan langs 2 wegen plaatsvinden:
- Extrinsieke inductie. Op de celmembraan bevinden zich receptoren (Fas-receptor, tumornecrosisfactor-receptor), die na binding aan een ligand een signaal aan de cel geven dat het apoptosemechanisme op gang brengt.
- Intrinsieke inductie. Apoptose kan van binnenuit de cel op gang komen, in geval van subletale beschadiging (chemotherapie, bestraling), bij verlies van cel-cel- of cel-matrixcontact of bij onvoldoende aanbod van groeifactoren, cytokinen of hormonen.
De effectorfase
In het celplasma is een aantal proënzymen aanwezig, die na een startsignaal elkaar in een specifieke volgorde activeren. De proënzymen betreffen 14 cysteïneafhankelijke aspartaatklievende proteasen, aangeduid als caspasen. Enkele daarvan, de initiatorcaspasen, slaan een lek in de mitochondriale membraan, waarna een aantal apoptosebevorderende eiwitten, onder andere cytochroom c, het celplasma binnenstroomt. Deze stoffen activeren de zogenaamde executorcaspasen. Deze caspasen splitsen een aantal structurele eiwitten, waarna de cel desintegreert.9
De gevoeligheid van een cel voor apoptose is afhankelijk van de mate waarin apoptosebevorderende en -remmende eiwitten tot expressie worden gebracht. Deze betreffen leden van de Bcl-2-eiwittenfamilie (de benaming stamt van het leukemisch B-cellymfoom, waarin dit eiwit voor het eerst werd geïdentificeerd). De Bcl-2-eiwitten hebben, afhankelijk van hun structuur, membraanstabiliserende (apoptoseremmende) of labiliserende (apoptosebevorderende) effecten.10 Een ander eiwit dat de gevoeligheid voor apoptose bepaalt, is p53, het product van het p53-tumorsuppressorgen. Het p53 blokkeert de voortgang van de celcyclus in geval van DNA-schade en biedt zo de tijd voor DNA-reparatie. Wanneer de reparatie niet totstandkomt, induceert p53 apoptose. Een mutatie in het p53-gen houdt in dat reparatie van DNA-schade niet plaatsvindt en dat de beschadigde cel niet wordt geëlimineerd. Voor de volledigheid zij vermeld dat de cel ook anticaspasen bevat, die in de toekomst mogelijk therapeutisch kunnen worden aangewend.11
De degradatiefase
Tijdens de degradatiefase treden veranderingen op in de membraan, in het celskelet en in de celkern.2 4 6 8 De cel schrompelt; het chromatine condenseert en het DNA wordt gefragmenteerd. Uiteindelijk blijven van de cel kleine blaasjes over, apoptotische lichaampjes, waarin de (afgebroken) celinhoud is opgesloten. Deze celresten worden door macrofagen en naburige cellen gefagocyteerd en komen niet in de weefsels terecht. Zodoende veroorzaakt apoptose, in tegenstelling tot necrose, geen lokale ontstekingsreactie en vindt men van de cel reeds enkele uren later niets meer terug.2
apoptose en ziekte
Normaal bestaat er een evenwicht tussen het aantal cellen dat wordt gevormd en het aantal dat in diezelfde tijd apoptotisch te gronde gaat. Aangezien de celproliferatie en -rijping enkele dagen tot weken in beslag nemen en de apoptose zich in enkele uren afspeelt, is in een evenwichtssituatie het aantal cellen, dat zich in apoptose bevindt, slechts een fractie (+-lymfocyten). Tijdens de ontwikkeling van het immunologisch systeem worden immunocyten, die lichaamseigen cellen zouden kunnen aanvallen, door middel van apoptose reeds in de thymus geëlimineerd. De celdodende werking van cytotoxische T-lymfocyten en ‘natural killer’-cellen (NK-cellen) berust op hun vermogen om in de doelwitcellen apoptose te induceren. Een aantal virussen weet te overleven doordat ze in de geïnfecteerde cel eiwitten tot expressie brengen die het apoptoseprogramma van de gastheercel blokkeren.
Een verstoring van het evenwicht tussen celgroei en -dood ligt ten grondslag aan het ontstaan van congenitale aandoeningen, van sommige maligniteiten, van auto-immuunziekten en van degeneratieve ziekten en ouderdomskwalen. Ziekten en aandoeningen waarbij ontregeling van het apoptoseprogram een rol speelt, zijn samengevat in tabel 1.
apoptose en prognose
Stamcellen hebben constitutioneel een hoge expressie van antiapoptotische en een lage expressie van proapoptotische eiwitten. Dit beschermt het organisme tegen ontijdig verlies van stamcellen. Dit fenomeen weerspiegelt zich in het gedrag van tumoren. De prognose van een maligniteit hangt samen met de gevoeligheid voor het ondergaan van apoptose. Hoe hoger de expressie van antiapoptotische eiwitten (Bcl-2-familie), des te minder gevoelig is de tumor voor cytotoxische behandeling en des te ongunstiger is de prognose.12-15 Hetzelfde geldt voor tumoren die een mutatie in het p53-gen hebben of autocriene groeifactoren produceren: ook deze tumoren kunnen moeilijk tot apoptose worden gebracht.16-19 Sommige tumoren brengen een namaak-Fas-receptor tot expressie en weten zo te ontsnappen aan de ‘doodsteek’ van het Fas-ligand, aanwezig op cytotoxische T-cellen en op NK-cellen.20 Er zijn ook tumoren die zich verdedigen tegen de afweer door zelf een Fas-ligand op hun celoppervlak tot expressie te brengen, waardoor ze Fas-positieve T-lymfocyten tot apoptose brengen zodra deze de tumor aanvallen.21
therapeutische modulatie van apoptose
Bevorderen van apoptose bij maligne aandoeningen
De oncolytische werking van chemo- en radiotherapie berust op inductie van apoptose in de tumorcellen. Niet altijd is het tumordodend effect van een cytotoxische behandeling voldoende omdat in sommige tumoren apoptose moeilijk op gang wordt gebracht, bijvoorbeeld omdat er een mutatie in het p53-gen bestaat of omdat de tumorcellen antiapoptotische eiwitten tot expressie brengen. Dergelijke oorzaken van therapieresistentie hoopt men te kunnen opheffen met moleculair-biologische manipulatie.
- Gentherapie. Met behulp van gentherapie kan men een stukje DNA dat codeert voor een receptor of een eiwit dat de neiging tot apoptose bevordert, via een vector aan de tumor aanbieden (transfectie). Wanneer vervolgens DNA-schade wordt toegebracht, gaan de getransfecteerde cellen alsnog in apoptose. Men gebruikt als vector een zogenaamd ‘replicatie-deficiënt’ RNA- of DNA-virus of liposomen waarin het betreffende DNA is ingebouwd. Men kan zo bijvoorbeeld een cel met een gemuteerd p53-gen voorzien van een normaal (zogenaamd ‘wild type’) p53 (figuur). Het moduleren van het apoptoseprogramma met behulp van gentherapie staat nog in de kinderschoenen (de Nederlandse Vereniging voor Gentherapie werd in 1999 opgericht). Hoewel deze therapie veelbelovend lijkt, is er nog een aantal obstakels. Zoals de transfectie thans gebeurt, blijft die niet beperkt tot de tumorcellen, maar worden ook andere weefsels met het gemanipuleerd virus geïnfecteerd en worden metastasen juist niet of onvoldoende bereikt. De voorlopige resultaten van enkele fase-I-/fase-II-trials met p53-transfectie bij patiënten met therapieresistente maligniteiten zijn samengevat in tabel 2.22-25 Onlangs is in de VS een met gentherapie behandelde patiënt overleden (overigens geen tumorpatiënt, maar een patiënt met een erfelijke stofwisselingsstoornis), zodat deze vorm van therapie voor deze indicatie door de Food and Drug Administration (FDA) tijdelijk is stopgezet.26
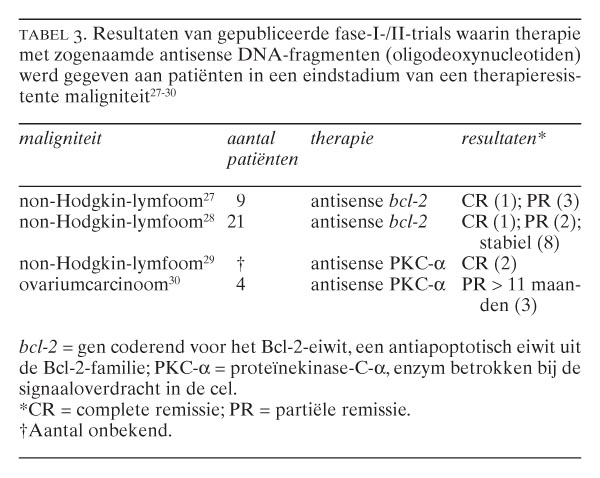
- Antisense DNA. Een andere benadering om het apoptotisch mechanisme te moduleren bestaat uit het toedienen van zogenaamd antisense DNA. Dit bestaat uit synthetische oligodeoxynucleotiden (ODN's), die complementair zijn aan het boodschapper-RNA van het eiwit, waarvan men de productie wil blokkeren. Als de ODN's de cel binnenkomen, hybridiseren ze met het betreffende boodschapper-RNA, waardoor de cel de informatie op het RNA niet meer in eiwit kan vertalen en het eiwit niet tot expressie komt (zie de figuur. Men kan zo bijvoorbeeld de expressie van de antiapoptotische Bcl-2-eiwitten blokkeren, waardoor de cel gevoeliger wordt voor cytotoxische behandeling.27 Een klinisch fase-I-onderzoek naar het effect van antisense bcl-2-ODN's bij patiënten in een eindstadium van non-Hodgkin-lymfoom resulteerde inderdaad in tumorregressie.27-30 Thans is een fase-II-onderzoek gestart om het therapeutische effect van antisense bcl-2-ODN's te testen in combinatie met chemotherapie.28 De eerste resultaten van deze klinische trials zijn samengevat in tabel 3. 27-30
Remmen van apoptose bij degeneratieve ziekten
Bij neurodegeneratieve aandoeningen, zoals de ziekte van Alzheimer of Parkinson bestaat een versneld verlies van neuronen, die apoptotisch te gronde gaan.31 Men heeft bij ratten en kippen met een vergelijkbare, erfelijke of toxisch opgewekte, degeneratie van het centrale zenuwstelsel, deze tot staan kunnen brengen door toediening van transformerende groeifactor-? (TGF-?).32 Recombinant humaan insulineachtige groeifactor (rhIGF-1) beïnvloedt het beloop van amyotrofische laterale sclerose gunstig doordat de apoptose van de voorhoorncellen wordt afgeremd.33 Lokale behandeling van neurotrofische keratopathie met zenuwgroeifactor remt het optreden van epitheeldefecten en bevordert de genezing van ulceratie en perforatie van de neuropathische cornea.34
conclusie
De nieuwe kennis over het biochemisch mechanisme van apoptose heeft geleid tot een nieuwe, zij het bescheiden, maar veelbelovende, therapeutische benadering van een aantal tot nu onbehandelbare ziekten. De eerste successen van moleculaire apoptosebevorderende therapie werden geboekt bij de behandeling van maligne aandoeningen. Het is te verwachten dat een aantal degeneratieve aandoeningen en ouderdomsziekten gunstig zullen reageren op apoptoseremmende interventies.
Literatuur
Kerr JFR, Wyllie AH, Currie AR. Apoptosis: a basicbiological phenomenon with wide-ranging implications in tissue kinetics. Br JCancer 1972;26:239-57.
Haanen C, Vermes I. Cel en ziekte. III. Apoptose, debiologische tegenhanger van mitose.Ned Tijdschr Geneeskd1993;137:1914-7.
Vaux DL, Haecker G, Strasser A. An evolutionaryperspective on apoptosis. Cell 1994;76:777-9.
Vermes I, Haanen C. Apoptosis and programmed cell death inhealth and disease. Adv Clin Chem 1994;31:177-246.
Leach AP. Apoptosis: molecular mechanism for physiologiccell death. Clin Lab Sci 1998;11:346-9.
Vermes I, Haanen C, Reutelingsperger CPM. Molecularbiology of apoptosis and programmed cell death. In: Aruoma O, Halliwell B,editors. Free radicals and molecular biology of human diseases. Berkshire,UK: Harwood Academic Publishers; 1998. p. 225-86.
Vaux DL, Korsmeyer SL. Cell death in development. Cell1999; 96:245-54.
Vermes I, Haanen C, Steffens-Nakken H, Reutelingsperger C.A novel assay for apoptosis. Flow cytometric detection of phosphatidylserineexpression on early apoptotic cells using fluorescein labelled Annexin V. JImmunol Methods 1995;184:39-51.
Saikumar P, Dong Z, Mikhailov V, Denton M, Weinberg JM,Venkatachalam MA. Apoptosis: definition, mechanisms, and relevance todisease. Am J Med 1999;107:489-506.
Mehmet H. Caspases find a new place to hide. Nature2000;403:29-30.
Yaoita H, Ogawa K, Maehara K, Maruyama Y. Attenuation ofischemia/reperfusion injury in rats by a caspase inhibitor. Circulation1998;97:276-81.
Miyashita T, Reed JC. Bcl-2 oncoprotein blockschemotherapy-induced apoptosis in a human leukemia cell line. Blood1993;81:151-7.
Dole M, Nunez G, Merchant AK, Maybaum J, Rode CK, BlochCA, et al. Bcl-2 inhibits chemotherapy-induced apoptosis in neuroblastoma.Cancer Res 1994;54:3253-9.
Campos L, Sabido O, Viallet A, Vasselon C, Guyotat D.Expression of apoptosis-controlling proteins in acute leukemia cells. LeukLymphoma 1999;33:499-509.
Kassim SK, Ali HS, Sallam MM, Fayed ST, Seada LS,abd-Elkawy E, et al. Increased bcl-2 expression is associated with primaryresistance to chemotherapy in human epithelial ovarian cancer. Clin Biochem1999;32:333-8.
Eliopoulos AG, Kerr DJ, Herod J, Hodgkins L, Krajewski S,Reed JC, et al. The control of apoptosis and drug resistance in ovariancancer: influence of p53 and Bcl-2. Oncogene 1995;11:1217-28.
Kurvinen K, Syrjänen K, Syrjänen S. p53 andbcl-2 proteins as prognostic markers in human papillomavirus-associatedcervical lesions. J Clin Oncol 1996;14:2120-30.
Perego P, Giarola M, Righetti SC, Supino R, Caserini C,Delia D, et al. Association between cisplatin resistance and mutation of p53gene and reduced bax expression in ovarian carcinoma cell systems. Cancer Res1996;56:556-62.
Ferreira CG, Tolis C, Giaccone G. p53 andchemosensitivity. Ann Oncol 1999;10:1011-21.
Pitti RM, Marsters SA, Lawrence DA, Roy M, Kischkel FC,Dowd P, et al. Genomic amplification of a decoy receptor for Fas ligand inlung and colon cancer. Nature 1998;396:699-703.
Hahne M, Rimoldi D, Schröter M, Romero P, SchreierM, French LE, et al. Melanoma cell expression of Fas(Apo-1/CD95) ligand:implications for tumor immune escape. Science 1996;274:1363-6.
Roth JA. Restoration of tumour suppressor gene expressionfor cancer. Forum (Genova) 1998;8:368-76.
Swisher SG, Roth JA, Nemunaitis J, Lawrence DD, Kemp BL,Carrasco CH, et al. Adenovirus-mediated p53 gene transfer in advancednon-small-cell lung cancer. J Natl Cancer Inst 1999;91:763-71.
Clayman GL, Frank DK, Bruso PA, Goepfert H.Adenovirus-mediated wild-type p53 gene transfer as a surgical adjuvant inadvanced head and neck cancers. Clin Cancer Res 1999;5:1715-22.
Habib NA, Ding SF, el-Masry R, Mitry RR, Honda K, MichailNE, et al. Preliminary report: the short-term effects of direct p53 DNAinjection in primary hepatocellular carcinomas. Cancer Detect Prev1996;20:103-7.
Smaglik P. Tighter watch urged on adenoviral vectors . .. Nature 1999;402:707.
Webb A, Cunningham D, Cotter F, Clarke PA, di Stefano F,Ross P, et al. Bcl-2 antisense therapy in patients with non-Hodgkin lymphoma.Lancet 1997;349:1137-41.
Cotter FE. Antisense therapy of hematologic malignancies.Semin Hematol 1999;36 (4 Suppl 6):9-14.
Nemunaitis J, Holmlund JT, Kraynak M, Richards D, BruceJ, Ognoskie N, et al. Phase I evaluation of ISIS 3521, an antisenseoligonucleotide to protein kinase C-alpha, in patients with advanced cancer.J Clin Oncol 1999;17:3586-95.
Yen AR, Halsey J, Fisher GA, Holmlund JT, Geary RS, KwohTJ, et al. Phase I study of an antisense oligonucleotide to protein kinaseC-alpha (ISIS 3521/CGP 64128A) in patients with cancer. Clin Cancer Res1999;5:3357-63.
Waggie KS, Kahle PJ, Tolwani RJ. Neurons and mechanismsof neuronal death in neurodegenerative diseases: a brief review. Lab Anim Sci1999;49:358-62.
Flanders KC, Ren RF, Lippa CF. Transforming growthfactor-betas in neurodegenerative disease. Cost effectiveness of recombinanthuman insulin-like growth factor-I therapy in patients with ALS. ProgNeurobiol 1998;54:71-85.
Ackerman SJ, Sullivan EM, Beusterien KM, Natter HM,Gelinas DF, Patrick DL. Cost effectiveness of recombinant human insulin-likegrowth factor I therapy in patients with ALS. Pharmacoeconomics1999;15:179-95.
Lambiase A, Rama P, Aloe L, Bonini S. Management ofneurotrophic keratopathy. Curr Opin Ophthalmol1999;10:270-6.
Artikelinformatie
Citeer dit artikel als



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Reacties